
A. M. Makarieva, V. G. Gorshkov
- Who is the Champion of Champions: Sergey Kirdyapkin, Ivan Ukhov, Usain Bolt or Michael Phelps?
- What would have happened to a flea had it jumped to a height of 2 meters?
- In which sports is man second only to the donkey and the elephant?
- Long-term disqualification of Homo sapiens on the Olympics of Life
All animals can move. But each species has perfected in a particular type of motion. Some animals set records in running, others are particularly good at walking or crawling, yet others are outstanding arboreal acrobats. Swifts fly better than anybody else, but they are pathetic on land where they can only crawl very slowly searching for a small hill from which they could take off. Loons fly very well both in the air and underwater, but they cannot walk. It is a huge problem for a loon to approach her nest moving on the ground – because of that the loon must build the nest very close to the water edge of a lake or a river (they cannot nest on the seashore because sea water comes and goes with tides). Moles can dig tunnels under the ground at a rate of up to 10 cm per second, but they practically never show up above the ground. Brachiating monkeys that rely on their hands to move in the tree canopy make great acrobatic performances on the trees but avoid coming down to the ground.
How are all these and other species-specific skills maintained? The answer comes from considering the stability principles of life organization. All animal species form populations of individuals. There are no species that would exist in the form of a single self-reproducing individual. The biological meaning of population is to ensure a continuous process of competitive interaction between individuals. This competitive interaction identifies individuals with various defects and removes such individuals from the population in one way or another. This is necessary because life is a super-organized process which complexity is unmatched in the inanimate world. This extraordinary high organization is prone to continuous decay. In order to sustain the high level of organization of the biological species it is necessary to remove individuals with defects from the population at a rate that exceeds the rate at which the genetic program of the species decays [1]. We note that the worst impairments of the genetic program concern the animal behavior. Individuals with behavioral defects (rapists, liars, cowards, fools etc.) experience the strongest selection pressure.

Loon’s nest
Sex is the most common and effective form of removing defective individuals from the population. Sex is realized via a complex system of powerful emotions and instincts. It is based on a division of all individuals in a population into two kinds: males and females. Males are unable to produce offspring. Females are able to produce offspring but only if allowed for by males. Strict competitive interaction between males makes it possible to identify defective males. Sexual claims of such males are inhibited in the population and they are excluded from the process of reproduction. Normal males that have withstood competition choose normal females to copulate with and do not interact with defective females. In this manner both defective males and defective females are barred from reproduction. But they can remain perfectly viable and retain the biological reproductive capacity. This allows the population to survive in natural catastrophes that can cause death of the majority of individuals in the population. Owing to the peculiarities of the recombination of parental genetic program during reproduction, the progeny of defective parents always comprises a small number of normal individuals. Therefore, competitive interaction of individuals and their sex-based selection greatly intensifies following major population catastrophes.
Under normal conditions the competitive interaction between females is much weaker than it is between males. This leaves more energy and time for females to produce and raise offspring. However, in periods of catastrophic overpopulation, when a rapid decline in population numbers is an ecological imperative, the competitive interaction between females who refrain from reproduction can become very intense, similar to competition between males. We also note that under normal conditions the share of defective individuals in the population is vanishingly small, such that competitive interaction mostly works to identify individuals with the best species-specific characteristics among normal individuals.
Thus, a major prerequisite for the super-organized complexity of life to remain stable is the continuous competitive interaction between individuals and the demonstration of the results of this competition to individuals of both sexes in the population. Obviously, competition should be first of all aimed to identify the best individuals in the most important activities of the species, in particular, its specialized ways of locomotion. Swifts should identify individuals who fly best, loons should identify individuals who both fly and swim best of all and so on. It makes no sense to range snakes by height of their jumps or to range moles by speed of their running on the ground surface.
Olympic games that are very popular with a huge number of people on Earth pursue the same goal of identifying the most competitive individuals in different types of locomotion. Olympic sports are carefully selected, and their number continuously grows with time. However, it is obvious that there is an informal ranking of the various Olympic sports. The mass-media allocate a disproportionate amount of time and space to highlight the records of Usain Bolt in sprinting or the outstanding performance of Michael Phelps in swimming. On the other hand, the number of gold, silver, bronze medals is used to rank countries for their sports performance. This is, however, done without analyzing the specific locomotive skills of Homo sapiens as a biological species. Let us try to scientifically analyze these skills and estimate the rank of various medal and the Olympic competitive capacity of various countries from that point.
As we shall see, in the animal world, man stands out as of one the best walkers and jumpers, a mediocre distance runner and one of the worst sprinters and swimmers [2-5]. Maximum sprinting speed in animals grows proportionally to the square root of the linear body size l of the animal, Fig. 1. The upper limit of maximum sprinting speeds and its dependence on body size l is determined by air drag. During running and other types of ground locomotion the animal experiences two types of resistance. The first one is the ground friction that does not depend on motion velocity u and is proportional to body weight mg, where m is body mass and g is acceleration of gravity. The second one is air drag that is proportional to the square of velocity u2. In order that both resistances had one and the same dimension, the air drag that is proportional to u2 should be summed with the ground friction in the form of a dimensionless combination of variables. There is only one relevant dimensionless combination: it is called Froude number and is equal to u2/gl, where l is linear body size defined as l = (m/ρ)1/2, ρ is body density that is close to density of liquid water. Froude number in all animal record-holders in sprinting is one and the same and is equal to 13. Therefore, maximum velocity umax grows as umax = (13 gl)1/2, Fig. 1.
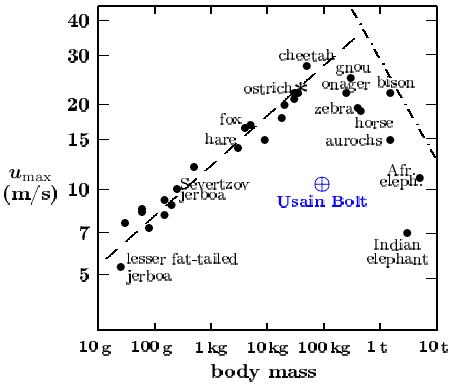
Fig. 1. Sprinting records in animals and man [2]. Dashed line umax = (13 gl)1/2 – maximum record speeds umax of animals with body mass not exceeding that of the cheetah. Dashed-dotted line umax = сl-1, c is a constant – the size-dependent limit on maximum speeds in larger animals imposed by the necessity to prevent overheating of the animal (that it would experience if develops a speed above the line).
Animals and birds maintain a constant body temperature. The metabolic power arising from food consumption is released within the body volume that is proportional to l3, while the resulting heat is dissipated to the environment via the body surface of area proportional to l2. The efficiency of transformation of total metabolic power to the power of locomotion is generally around 25% in animals, but rises to 40% during jumping. Ground friction being proportional to body weight, i.e. l3, the power of overcoming this friction is proportional to ul3. This power should be replenished by the power of food consumption that is proportional to l2. Therefore, if the air drag is smaller or equal to the ground friction, the maximum running velocity should decrease with growing body size as u=c/l (cl2 = ul3, where c is a constant).
The dash-dotted line in Fig. 1 corresponds to the equality between the total animal power spent on locomotion and the power of ground friction. Running at velocities above the dash-dotted line is not possible because of the necessity to maintain a constant body temperature (running at higher speeds will cause overheating). The dashed line in Fig. 1 corresponds to the equality between the power of air drag and the power of ground friction. Locomotion with speeds above the dashed line is possible but ineffective: as velocity increases, the air drag grows as the square of velocity and quickly exceeds the ground friction, which becomes negligibly small. As the contact with the Earth is minimized, running becomes impossible. It is at this point that transition occurs from running to flight, i.e. to the motion without any contact with the ground. Therefore, all winged flying animals are characterized by Froude number exceeding the magic number of 13, the threshold emphasized by nature itself. Thus, running speeds of animal record-holders with body mass not exceeding that of cheetah cluster around the dashed line in Fig. 1 with Froude number equal to 13. Larger animal runners have to cluster below the dash-dotted line. Finally, those animals for whom running is not the main thing in their life, reside well below both the dashed and the dash-dotted lines, Fig. 1.
Animals that run more slowly than the record-holders in their body size interval experience only ground friction. Air drag is insignificant for such animals because of low velocity. Among such sluggish animals the most rapid will be those who have the longest strides and thus touch the ground fewer times per unit distance. This explains the world records of Usain Bolt who has a large body mass (large l) and long strides. The majority of animals do not need maximum sprinting speeds and they are unable to develop them. Man belongs to this majority – the record sprinting velocity in man coincides with that of a 250 g jerboa. Meanwhile the record sprinting velocity of a 50 kg cheetah is three times higher than Usain Bolt’s Olympic and world records, Fig. 2. Among animals of the same size as Homo sapiens none performs worse in sprinting than man does. The same can be said about swimming. Dolphins, seals and many other animals swim many times faster than Michael Phelps, the legendary American sportsman.
Nonetheless as a jumper and walker our species ranks very high in the animal world. Maximum jump height does not depend on body size starting from the body size of galago, who has a body mass of 200 g and is able to display a standing jump of up to 230 cm. With just a small run-up the best animal hoppers ranging in body mass from 200 g to 400 kg jump to a height of 200-240 cm, Fig. 2. The London Olympics achievements of Russian height jumpers Ivan Ukhov (238 cm) and Anna Chicherova (205 cm) are in no way less spectacular than the records of the best animal jumpers.
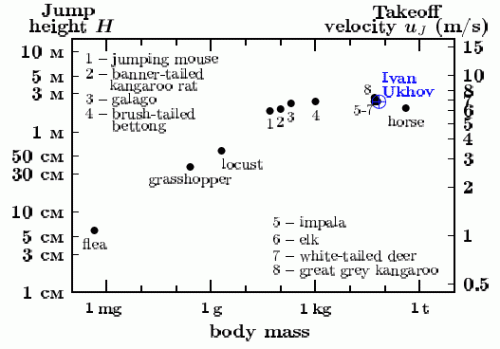
Fig. 2. High jumping in distinguished animal hoppers and man [2]
In pole vault, a discipline that is not practiced by any other animal except Homo sapiens, the same unique abilities of our species are manifested. The takeoff velocity during the pole vault is determined not only by the muscle impulse during the takeoff, but also, and mostly, by the velocity of the approach run. Via the elastic deformation of the pole the kinetic energy of the approach run and takeoff is transformed into the potential energy of ascent. It is this potential energy that together with the additional effort of the arm muscles pulling the body up the pole, determines the ultimate height H of the pole vault. The same energy of approach run and takeoff determines distance L of the long jump. Neglecting the pull-up of the pole vaulter, height H of the pole vault and length L of the long jump are connected by a simple physical relationship, L = 2H. The legendary Soviet sportsman Sergei Bubka remains an unrivaled world record-holder in male pole vaulting, while Russian sportswoman Yelena Isinbayeva is a holder of 28 world records in female pole vaulting.
Note that the maximum jumping height of small animals with body mass not exceeding 100 g decreasing with diminishing body mass. Thus, for example, a 0.25 g cricket jumps up to 20 cm, while a 0.45 mg flea jumps up to 6 cm (and not up to 2 m contrary to folklore), Fig. 2. During the jump the biochemical energy is released that has been previously accumulated in the animal body. Jump height is determined by the equality of the kinetic and potential energy, muJ2/2 = mgH (the energy conservation law). Thus this height does not depend on animal body mass: H = uJ2/2g, where uJ is the vertical takeoff velocity. Locomotive power per unit body mass developed by the animal as it jumps is related to velocity u and linear body size l. It should be proportional to u3/l, as it is the only combination with the dimension of mass-specific power that can be built with help of two variables u and l. At a given value of takeoff velocity uJ determined from the energy conservation law, the power of the jump increases with diminishing body size l and reaches the biochemical limit of qmax = 5 W/g for animals sized as the Senegal galago (m ~ 200 g), Fig. 2. Any further increase in jumping power with decreasing body size l is biochemically prohibited: the maximum jump height is limited by the condition uJ3/l = qmax. In other words, uJ = (qmaxl)1/3, which means that for the smaller animals jump height H is proportional to l2/3. In the result, the smaller hoppers like locusts, crickets and fleas, can jump to a much lower height than the larger animals, Fig. 2.
In order for a flea with body size l ~ 1 mm to hop over a 2.38 m bar, to rival the London Olympic champion Ivan Ukhov, the flea would have had to develop such a high metabolic power that the life biochemical machinery cannot endure. Per unit mass, this power would constitute about u3/l ~ 200 W/g >> qmax. It would have been the last jump in the flea’s life, as the flea would have simply burnt when rocketing to over 2 m height.
Long and fast walking demands the organism to be maximally efficient in transforming the energy stores of the body into the kinetic energy of locomotion. We can define the mean daily velocity of locomotion u0, which is proportional to the energetic efficiency of locomotion (it is similar to how much a car spends to move one kilogram of its mass over a hundred kilometers). This velocity does not depend on body mass and varies within relatively universal taxon-specific limits in all animals, Fig. 3. By this parameter, a Homo sapiens walker ranks close to the maximum of the entire animal world, Fig. 3. Man-walker loses only to donkey and elephant. The London 2012 results of Russian race walkers Sergey Kirdyapkin (Olympic record, 3 hours 36 minutes in 50 km race, mean speed u = 3.9 m/s = 14 km/hour) and Elena Lashmanova (world record, 1 hour 25 minutes in 20 km race, the same mean speed u = 3.9 m/s = 14 km/hour) are record achievements in exactly that type of locomotion where Homo sapiens is one of the best specialists. In other words, it is in this field that human body functions at a high world standard common to all life. (Note however that namely this extraordinary ability to walk fast and long made it possible for humans to invade all land ecosystems and ultimately bring them to the modern miserable state. The African elephant has likewise possibly contributed to the large-scale deterioration of plant cover on the African continent. It is only donkey who, being an equally efficient walker, has not in any way undermined the stability of its natural habitat.)

Fig. 3. Mean daily velocity of locomotion u0 maintained during adult life in animals and man [2,4]. The higher the energetic efficiency of locomotion, the higher u0.
Wrestling is widely spread among males in the animal world. Here too man is among the best performers, but does not exceed the pan-life standards. For example, male vipers in spring exhibit breathtaking performances in “Greko-Roman wrestling”. During the fight, the sportsvipers show noble mutual respect and never use their poisonous bite against the contender.
Individuals in human population need and have to be able to walk, to throw stones and to jump – these are genetically programmed capabilities of Homo sapiens. Like many other animal species, e.g., bats or moles, our species did not need to run or swim fast. As a consequence, humans run and swim conspicuously more slowly and inefficiently than do animal species specializing in these types of locomotion. Therefore, human champions in sprinting and swimming claim extraordinary attention of the general public. Meanwhile these records would not impress individuals of other species if the latter were invited as referees. It is difficult to imagine a tropical arboreal monkey deeply impressed by the Olympic gymnastics or a solan-goose applauding Olympic divers (solans are the bird divers able to enter water without splash at a speed of over 100 km per hour).

Spring Greko-Roman wrestling of two male vipers
However, no species in the animal world are able to throw objects as far as humans do. No species play tennis, volleyball or basketball. These and other sporting games do not only involve unique types of locomotion but also require rapid thinking and decision-making. Gold medals of Tatyana Lysenko, Russian Olympic record-holder in hammer throw, as well as the dramatic victories of the Russian volley-ball (gold) and basketball (bronze) teams during the last days of the Olympic games in London stand out of the other remarkable Olympic results as they rank among records of life as a whole.
To summarize, the achievements of our country in London 2012 are special. They could have been applauded by all life on the planet – had it not been for the long-term disqualification of Homo sapiens for his unsporting behavior with respect to life’s other sportsmen, whose natural habitats humans have been most actively destroying.
Literature
[1] Gorshkov V.G., Makar’eva A.M. (2001) On the possibility of physical self-organization of biological and ecological systems. Doklady Biological Sciences, 378, 258-261.
[2] Gorshkov V.G. (1983) Power and rate of locomotion in animals of different sizes. Journal of General Biology, 44(5), 661-678.
[3] Gorshkov V.G. (1984) Energetical efficiency of flight and swimming. Journal of General Biology, 45, 779-795.
[4] Gorshkov V.G., Gorshkov V.V., Makarieva A.M. (2000) Biotic regulation of the environment: Key issue of global change. Springer-Praxis, London.
[5] Gorshkov V.G. (1995) Physical and biological bases of life stability. Man. Biota. Environment. Springer, Berlin.

Pingback: К итогам Лондонской Олимпиады: энергетические рекорды людей и животных | Наука изнутри